在寒武纪,有种比其他动物要大若干倍的动物,那就是射齿目,俗称“奇虾”。它是寒武纪时期海洋生态系统中的顶级捕食者。
它们长有一对巨大坚硬的用来捕食的前附肢,巨大的柄状复眼以及身体两侧一系列用来游泳的桨状叶片(图1)。

图1 射齿目典型代表(图片来自网络)
我们采集到的射齿目化石有至少90%都只能看到它们的前附肢,而身体其它部分很难在化石上保存下来。这是因为前附肢非常坚硬,而它的身体异常柔软,因此全世界保存有完整身体的射齿目化石更是凤毛麟角。基于这种条件,大部分情况下古生物学家都只能依靠前附肢的形态特征来建立新的属种。
目前,射齿目可划分为4个科,分别是奇虾科、抱怪虫科、筛虾科以及赫德虾科(图2)。有超过20个属25个种被报道,它们分布在北美、中国、欧洲、澳大利亚、非洲以及格陵兰等地的28个特异埋藏化石库中。其中以华南的澄江生物群、加拿大的布尔吉斯页岩生物群以及美国犹他州Wheeler组中的射齿目多样性最大。
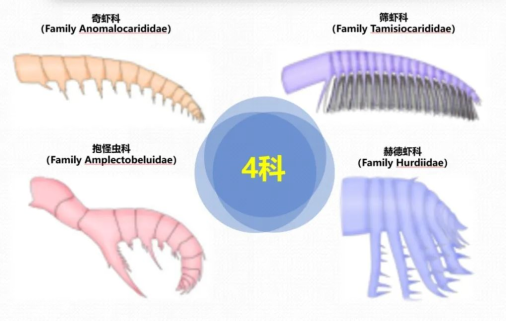
图2 射齿目4个科以及前附肢形态
01射齿目前附肢解剖学构造
射齿目的前附肢可见很多分节,由两部分构成,一部分是靠近身体部分的基部,另外一部分是离身体较远的螯部,两部分经常会形成一定的角度(θ)。螯部每一肢节内侧会长有刺状构造(内叶),不同类群会有不同的形态。而刺的前后边缘也会生长不同数量的小刺(辅助刺)。而爪子的外边缘也会生长数量不等的背刺(见图3)。
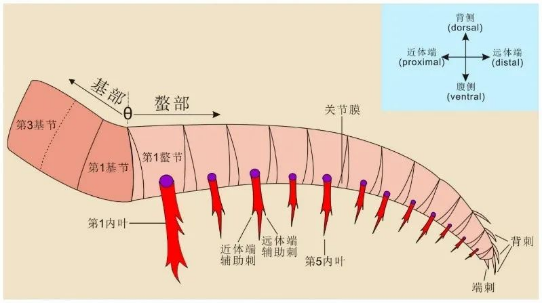
图3 射齿目前附肢解剖学构造示意图(引自吴雨,2021)
可以看到,奇虾科的前附肢细长,内叶较短,辅助刺较少;抱怪虫科则有一根过度膨大的内叶;筛虾科的内叶较长,并且辅助刺十分密集;赫德虾科的内叶呈长的刀片状,并且辅助刺只分布在一侧边缘。
02澄江生物群射齿目新属种
中国云南的澄江生物群是了解寒武纪大爆发的重要窗口,其物种多样性以及化石保存的精美程度不亚于加拿大布尔吉斯页岩生物群。在澄江生物群中已经报道了许多射齿目类群,如Amplectobelua symbrachiata、Lamincaris chimera、Lyrarapax unguispinus、Ly. trilobus、Ramskoeldia platyacantha 和R. consimilis。但是,这些类群仅限于抱怪虫科,对于其它科的报道几乎没有。
近期,西北大学早期生命研究团队的吴雨博士等人报道了澄江生物群中射齿目奇虾科和筛虾科的代表属种,分别为新种狼牙光滑虾Lenisicaris lupata,加拿大奇虾相似种以及新的融和种帚状侯氏虾Houcaris saron (Hou, Bergström and Ahlberg, 1995) comb. nov.。
2.1 奇虾科
狼牙光滑虾Lenisicaris lupata的前附肢细长,发育3个基节和13个螯节;第1螯节至第12螯节各发育一对内叶,内叶两侧边缘光滑不发育辅助刺;奇数螯节上的内叶长度大于偶数螯节;内叶长度与所在螯节的高度近似相等,向远体端逐渐变短;第10螯节至第12螯节背边缘各发育一根背刺;第13螯节发育一根背刺和一根端刺(图4,5)。此属种代表了澄江生物群中第1个正式命名的奇虾科类群。
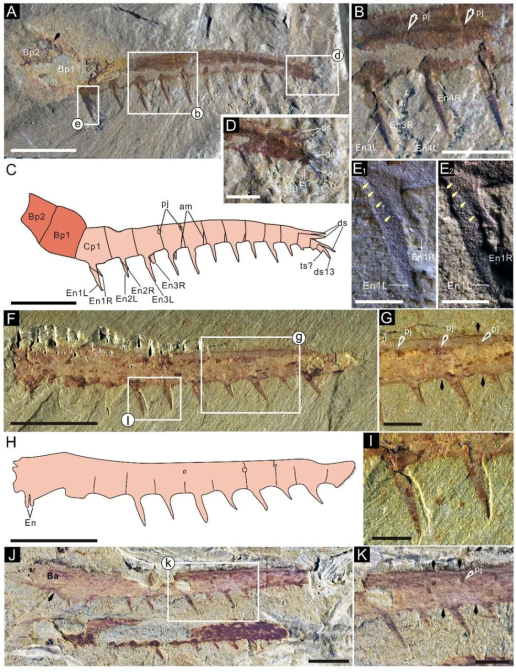
图4 澄江生物群中 Lenisicaris lupata前附肢标本

图5 Lenisicaris lupata前附肢复原图
加拿大奇虾近似种Anomalocaris cf. canadensis Whiteaves, 1892与奇虾模式种Anomalocaris canadensis极为相似,它们都发育有较短的内叶以及内叶两侧一对辅助刺(图6)。
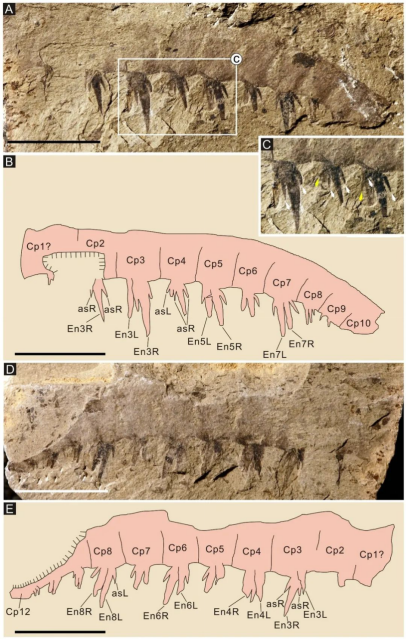
图6 澄江生物群中Anomalocaris cf. canadensis Whiteaves, 1892前附肢
2.2 筛虾科
帚状侯氏虾Houcaris saron (Hou, Bergström and Ahlberg, 1995) comb. nov.:Houcaris saron前附肢共16个肢节,包括3个基节和13个螯节,第2螯节至第8螯节近于正方形,第1内叶呈刀片状并向远腹端弯曲;第2内叶至第12内叶呈拉长杆状,其长度是所在螯节高度的1.7倍;内叶长短交替排列,其奇数螯节上的长于偶数螯节上的;第2内叶至第12内叶发育两根后辅助刺;第2内叶至第8内叶发育5根前辅助刺,第9内叶至第11内叶则发育2根前辅助刺(图7)。
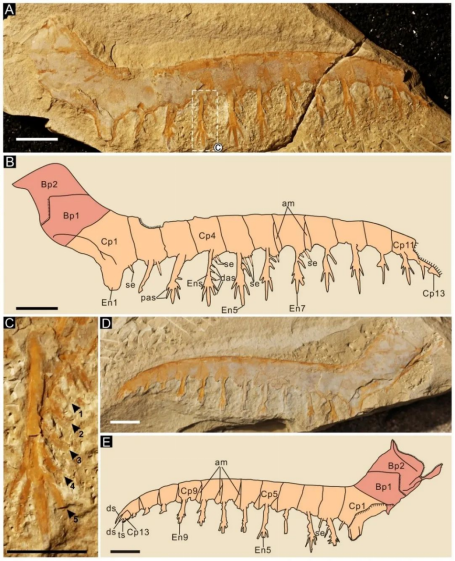
图7 澄江生物群中Houcaris saron (Hou, Bergström and Ahlberg, 1995) comb. nov.前附肢
最初被侯先光将其命名为帚状奇虾Anomalocaris saron,并被放到奇虾科中。但是因为其前附肢发育修长的内叶以及较多且长的辅助刺,本研究为其建立新属侯氏虾属Houcaris,并将其放入到筛虾科中。因此,帚状侯氏虾代表华南首次报道的筛虾科类群。
03奇虾科与筛虾科埋藏环境
本研究基于奇虾科和筛虾科所有类群在各个代表不同沉积环境的特异埋藏化石库中的分布来推测它们当时的生活环境和埋藏环境。
3.1 奇虾科
奇虾科在不同特异埋藏化石库中的分布表明此类群能够生存于各种不同的海水环境当中,从陆架内缘到外缘,再到碳酸盐台地边缘。Lenisicaris的两个种以及Anomalocaris的绝大多数种通常与陆架外缘沉积环境相关,典型的以华南的澄江生物群和凯里生物群以及美国宾夕法尼亚州Kinzers组为代表。一般认为,澄江生物群的沉积环境位于远离滨岸的陆架外缘,并且其中的化石只经过了极短距离的搬运。
我们的化石材料均采集于澄江-海口-安宁地区,此地区中的化石被认为是原地(autochthonous)或近原地(parautochthonous)埋藏,表明这些化石能够反映原始群落的真实特征。因此本文认为其沉积环境可近似代表当时奇虾科的生活环境。Anomalocaris也存在于湖南杷榔组以及布尔吉斯页岩生物群。湖南杷榔组代表较深水斜坡的沉积环境,布尔吉斯页岩生物群被解释为靠近斜坡的碳酸盐岩台地边缘环境(Wu et al., 2021a)。3.2 筛虾科
筛虾科在特异埋藏化石库中的分布表明此类群更加偏好于浅水且含氧状况良好的环境。Houcaris magnabasis存在于内陆架至陆架中部的环境,其中内陆架以美国内华达州Pioche组为代表,Carrara组则代表陆架中部的环境。H. saron 仅见于澄江生物群中,被普遍认为代表较浅的碎屑岩台地外缘环境,并且其中的化石为原地埋藏或近原地埋藏。
南澳大利亚鸸鹋湾页岩中的Anomalocaris briggsi Nedin, 1995表明筛虾科也能够生活在近岸带更浅水的环境中。Tamisiocaris 似乎是一个特例,因为T. borealis 存在于代表残存台地外边缘环境的Sirius Passet 生物群中,其相似种Tamisiocaris aff. borealis 存在于Kinzers 组Fine Pelitic Facies of the Emigsville 段,代表间歇脉冲沉积(intermittent, pulsed sedimentation)的低能深水环境。但是这两个沉积区域中的沉积物被认为经过了远距离搬运而来的。这表明沉积区的生物体包括筛虾科被埋藏在远离生存环境的地方。
也就是说,Tamisiocaris 也很可能生存在浅水透光带环境中。事实上,筛虾科一般被认为是滤食性动物,而深水低氧的环境不利于初级生产者和浮游生物的生存,无法为其提供充足的食物来源(Wu et al.,2021b)。
04奇虾科与筛虾科生物古地理模式
4.1 奇虾科
奇虾科时代延限为寒武纪第3期至古丈期,分布于华南(云南)、劳伦和东冈瓦纳(南澳大利亚)地区(图8)。Anomalocaris属于全球属,其广泛的分布很可能与其浮浪幼虫在海水中的扩散有关。奇虾科的分布范围局限于低纬度地区,表明其分布很可能受到气候带和温度的控制。古丈期的Weeks生物群与其它劳伦大陆上的第3世生物群在生物组成上有重大的差别,表明古丈期的动物群落结构发生了明显的重构。这一结果无法简单地用水深、古地理或地层学来解释。相反,这代表了一次真正的生物演化事件,被认为是生命史上一个重要过渡时期的开始,将早古生代的两次主要的辐射事件(寒武纪大爆发和奥陶纪大幅射)分开。这个时期同时发生了全球三叶虫大灭绝事件。
而引起这次灭绝事件的原因包括:①海水的冷却导致了海洋的倾覆;②晚寒武世缺氧海水的周期性扩张,而这次缺氧事件得到了明显的δ13C正漂(Steptoean Positive Isotope Carbon Excursion, SPICE;Gerhardt and Gill,2016)和相关的δ34S正漂证据的支持。奇虾科的灭绝与这些因素也息息相关,同时作为三叶虫的捕食者,奇虾类也失去了食物资源而灭绝(Wu et al., 2021a)。
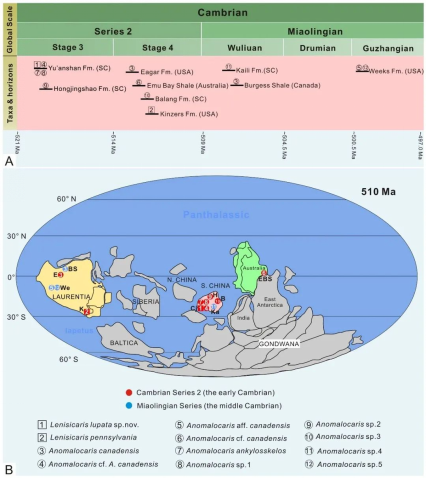
图8 奇虾科时空分布
4.2 筛虾科
筛虾科的化石记录目前仅见于寒武系第二统,分布时限较短,但地理分布较广泛,分布于华南(云南)、劳伦(美国和格陵兰岛)和东冈瓦纳(南澳大利亚)地区(图9)。华南澄江生物群中的Houcaris saron和北格陵兰Sirius Passet生物群中的Tamisiocaris borealis。同时代表最早的筛虾科化石记录,表明筛虾科从寒武纪第3期开始出现。在之后的第4期,筛虾科化石记录依然较少,分布于劳伦古陆西边缘的内华达州和宾夕法尼亚州,以及东冈瓦纳的南澳大利亚地区。最终,筛虾科化石记录消失在苗岭统以及更晚的地层中,这似乎代表真正的演化缺失,因为射齿目其它类群在苗岭世时期多样性以及丰度都很高。筛虾科在寒武纪第4期末的灭绝可能是受到了引起古杯,以及莱德利基虫和古油栉虫灭绝事件因素的影响,比如火山活动或海平面升降引起的缺氧海水的扩张,以及N元素损失和P输入增加导致的海水环境低营养化。筛虾科的分布范围也局限于低纬度地区,表明其分布很可能受到气候带和温度的控制(Wu et al., 2021b)。
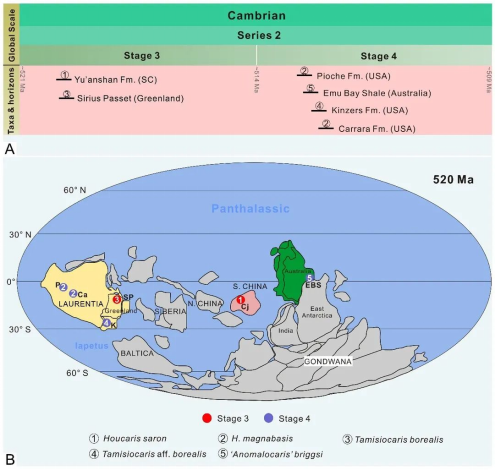
图9 筛虾科时空分布
本项研究由国家自然科学基金、中国科学院战略性重点研究计划和111项目共同资助。
论文相关信息:
Wu Y., Ma J.X., Lin W.L., Sun A., Zhang X.L. and Fu D.J. 2021a. New anomalocaridids (Panarthropoda: Radiodonta) from the lower Cambrian Chengjiang Lagerstätte: Biostratigraphic and paleobiogeographic implications[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 569:110333.
Wu Y., Fu D.J., Ma J.X., Lin W.L., Sun A., Zhang X.L. 2021b. Houcaris gen. nov. from the early Cambrian (Stage 3) Chengjiang Lagerstätte expanded the palaeogeographical distribution of tamisiocaridids (Panarthropoda: Radiodonta). PalZ, 2021b, 95: 209–221. doi.org/10.1007/s12542-020-00545-4.
吴雨.寒武纪澄江生物群射齿目节肢动物研究【D】.西北大学,2021.